Now that you have learned all about a single neuron, imagine a community of neurons. Individual neurons make connections to target neurons and stimulate or inhibit their activity, forming circuits that can process incoming information and carry out a response.
In this part you will learn about how neurons communicate and send signals in order to have an action or a response.
1) Signal transmission
Neurons communicate with one another at junctions called synapses. Synapses are usually between an axon and a dendrite of another neuron but there are different types of synapses as you can see in the figure 2.1.
In a neuron synapse we can find the presynaptic, or sending neuron, that causes the transmission of a signal from axon terminal to dendrite of another neuron, the postsynaptic, or receiving neuron.

Depending on the type of the stimulator, we can define two types of synapses: electrical and chemical synapses.
a) Chemical synapses
Chemical transmission involves a release of chemical messengers known as neurotransmitters. Neurotransmitters carry information from the presynaptic neuron to the postsynaptic neuron.
Inside the axon terminal of a sending cell, there are many synaptic vesicles. These are membrane-bound spheres filled with neurotransmitter molecules. There is a small gap between the axon terminal of the presynaptic neuron and the membrane of the postsynaptic cell, and this gap is called the synaptic cleft.
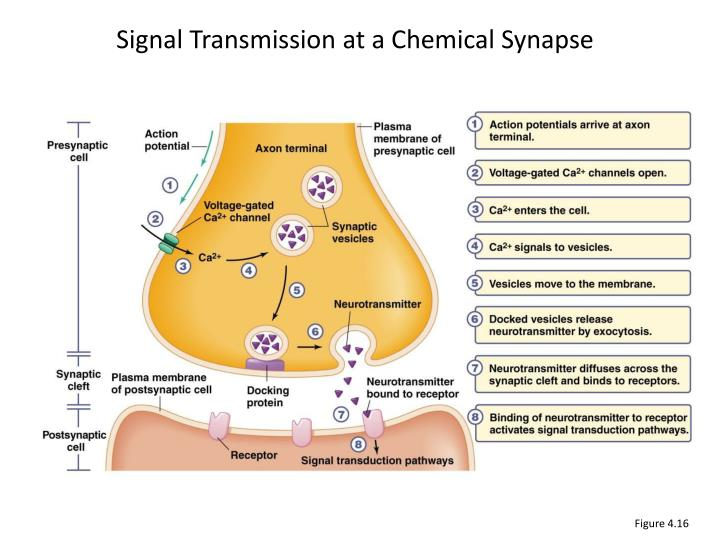
Now that you know the definitions let’s see how the signal is transmitted in a chemical synapse.
First, the action potential or nerve impulse arrives at the axon terminal (synaptic terminal) of a presynaptic neuron. The primary function of the synaptic terminal is to propagate action potentials from the axon to the presynaptic membrane. Recall that propagation of an action potential requires the presence of both voltage-gated Na+ and K+ channels.
The depolarization of the neuron will activate the voltage-gated calcium channels in the cell membrane. The electrical and chemical gradients for Ca2+ ions leads the Ca2+ ions to flow inside the neuron (calcium is present at a much higher concentration outside the neuron than inside). Therefore the calcium rushes into the cell.
The Ca2+ allows synaptic vesicles to fuse with the docking protein at the axon terminal membrane and to release the neurotransmitters into the synaptic cleft by exocytosis.
Neurotransmitters then diffuse across the synaptic-cleft and bind to the receptors or ligand-gated channels of the postsynaptic cell. The activation of this membrane receptors leads to a change in opening or closing ion membrane channels. These effects on channels behavior can be direct when the receptor is a ligand-gated ion channel, or in other cases, the receptor is not an ion channel itself but activates ion channels through a signaling pathway.
This effect in behavior of channels can be in two types depending on the ions involved:
–Depolarization makes interior of the cell more positive.
–Hyperpolarization makes interior of the cell more negative than its resting value.
After the release of the neurotransmitters in the synaptic cleft, they fix themselves on the receptors of the postsynaptic neuron, as we said above.
However, the neurotransmitters need to be deactivated or removed. If not, there is a continuous activation and action potential firing on the postsynaptic neuron.
There are two main mechanisms of neurotransmitter deactivation: enzyme degradation and reuptake.
— The first mechanism, enzyme degradation can be typically applied to the neurotransmitter acetylcholine. When acetylcholine binds to the postsynaptic neuron receptor and depolarizes the cell (this mechanism will be explained later in part 2.2A of this chapter), it quickly unbinds the receptors to bind to the enzyme acetylcholinesterase. This enzyme dissociates the acetate and choline that form acetylcholine by breaking the ester bond which joins these two compounds. Acetylcholine is thus deactivated and cannot bind anymore to the receptors of the postsynaptic neuron.
— The second mechanism could be assimilated to a recycling process: the neurotransmitters are removed from the synaptic cleft with transporter proteins that take them back to the presynaptic terminal where they are packaged again in the vesicles. One of these neurotransmitters that are recycled in this way is serotonin. There is indeed the serotonin transporter called SERT which is a sodium, potassium and chloride dependent receptor.
The SERT first binds a sodium ion. Then, serotonin under its protonated form (5HT+) binds to the SERT and finally a chloride ions binds too. It leads to a change in the conformation of the transporter protein, which can cross the membrane of the presynaptic neuron to release the ions and the serotonin in the intracellular space. It then binds to a potassium ion that allows it to go out back in the extracellular space.
b) Electrical synapses
In electrical synapses, unlike chemical synapses, the two neurons are in physical connection. This connection between the presynaptic neuron and postsynaptic neuron is in a form of channel called gap junction, that allows the current-ions flow direct throw them.
In an electrical synapse the two cells can exchange cytoplasm therefore the transmission of signals is quicker, and can occur in both directions.
This advantage is especially obvious in neuronal pathways that serve as escape mechanisms, such as those in the tail muscles of crayfish and lobsters and those in the pectoral fins of fishes. In these cases, the rapidity of the animal’s response to imminent danger has a selective value.
Also, electrical synapses contribute to the synchronized activity of groups of cells. In many cases, they can carry current in both directions so that the depolarization of a postsynaptic neuron will lead to the depolarization of a presynaptic neuron.

Some synapses can be both electrical and chemical at these synapses the transmission as an electrical signal is more rapid.
2) Neurotransmitters, receptors and vesicles
As we saw earlier, the main function of neurotransmitters is being a chemical information carrier between the pre and postsynaptic neuron. In order to do so, most of these neurotransmitters have to be in vesicles.
There is more than 100 neurotransmitters types in the organism but we will see here only their main classes.
We can consider five different types of neurotransmitters which are:
● Acetylcholine
● Amino acids
● Biogenic amines
● Neuropeptides
● Gas
a) Acetylcholine
These neurotransmitters can be found in a neuromuscular junction and will mainly create excitatory postsynaptic potential. In this case, it will help the muscular contraction.
After a certain amount of time (when the organism don’t need more acetylcholine and all receptors are already linked) which is the end of the synaptic transmission, an enzyme called acetylcholinesterase is released into the synaptic cleft in order to hydrolyze acetylcholine.
There are two types of acetylcholine receptors:
Ionotropic receptors look like ion channels pores and lead to an excitatory postsynaptic potential when linked with acetylcholine (note that when you smoke, nicotine binds to these receptors).
Metabotropic receptors are indirectly linked with ion channels and need to interact with a second messenger in order to activate or inactivate those channels. Metabotropic acetylcholine receptors can be found in hearth muscular cells and will this time, through a second messenger called G protein, open K+ channels and lower the myocardial contraction frequency. In this case, acetylcholine will have an inhibitory effect.
b) Amino acids
The main amino acid neurotransmitter type in the central neural system is the glutamic acid which is an excitatory neurotransmitter that plays a role in the long term memory.
Another well known amino acid neurotransmitter is the gamma-aminobutyric (also called GABA) which have an inhibitory effect on brain synapses by increasing the membrane permeability to Cl- (you will see ions effect on the action potential in part 3).
c) Biogenic amines
Biogenic amines are neurotransmitters that come from amino acids and that are mainly composed of the three catecholamines (biogenic amines that derived from tyrosine): noradrenaline, dopamine, epinephrine (also called adrenaline but it is outweighed by its chemical precursor noradrenaline as a neurotransmitter in volume and importance), histamine and serotonin. Noradrenaline is an excitatory neurotransmitter that can be found in the autonomous neural system and that plays major roles in tonic and reflexive changes in cardiovascular tone (note that noradrenaline is also a hormone (see chapter “Hormones and endocrine system” in some cases)).
Glycine is the equivalent of gamma-aminobutyric neurotransmitters but out of the brain.
The central histamine system is involved in many brain functions such as arousal, control of pituitary hormone secretion, suppression of eating and cognitive functions.
Dopamine is a neurotransmitter that can be found almost everywhere in the brain, and this special molecule also affects sleep, humor, attention and learning.
Note that many addictive drugs increase dopamine release or block its reuptake into neurons after the release. These drugs can be addictive by increasing the dopamine level leading to a feeling of happiness spread into the body or also because of the long term potentiation (that will be explain in the next chapter).
d) Neuropeptides
This class is composed of short amino acids chains that link to metabotropic receptors to create a synaptic response.
One of those neurotransmitters is called substance P and leads to an excitatory response when linked to its receptor that increase the pain feeling.
Another neuropeptide, endorphine, is almost the opposite of the previous one.
This neurotransmitter is created by the brain in intense stress cases. It can be considered as a natural analgesic by creating an inhibitory response, that will lower the pain as well as create some other psychotic reactions (such as euphoria or respiratory slowing).
e) Gas
This class of neurotransmitters is a bit different, as gas cannot be maintained in vesicles. The consequence of it is the fact that gas should be synthesised “on demand” by cells.
Gas neurotransmitters are mainly nitric oxide (or nitrogen monoxide) that signal the surrounding smooth muscles to relax, by stimulating a cell’s membrane enzyme that then synthesize a chemical second messenger. It finally leads to a synaptic response. It plays a major role in the sexual excitation for men as it provokes a relaxation of erectile tissues (vasodilatation) that lead the corpus spongiosum to be blood-filled.
Another neurotransmitter gas is the carbon monoxide (created in really small quantities by vertebrate cells) that plays a role in the release of some hypothalamic hormones in the brain.
3) Action potential, depolarization, repolarization, hyperpolarization
As we already said before, there is a membrane potential inside neurons which is created by ions flows through selective ionic channels. Indeed, these channels can either be opened or closed by reacting to a stimulus. The channels state (open or close) determine the membrane permeability to certain type of ions.
We will now look deeply into the action potential creation mechanism.
a) The action potential creation
1) Resting phase:
At first, we consider that the membrane potential is in its resting state (around -70 mV). In this state, most of the sodium voltage-gated channels are closed (they only open at a certain membrane potential value which is higher than -70mV) as well as potassium voltage-gated channels that are mainly in a closed state. Note that these voltage gated channels do not work like previously seen leakage channels, they only open once an action potential is triggered. We can define three types of gates for voltage-gated channels:
– The gate m which is an activation gate that is normally closed, and opens quickly when the membrane potential becomes more positive (these gates are found in sodium channels)
– The gate h which is a deactivation gate that is normally opened, and closes when the membrane potential becomes too positive (these gates are found in sodium channels and contribute to the inactivation loop that we will see just after).
– Finally, the gate n which is normally closed, and slowly starts to open when the membrane potential becomes too positive (these gates are found in potassium channels)
Note that in the resting state, voltage-gated sodium channels are said deactivated (meaning that the m gate is closed but the h gate is open) and ions cannot flow through the membrane as one of the enter is closed.

2) Stimuli and depolarization:
Now, a stimulus (created by signal transduction from sensitive neurons) arrives and opens some voltage-gated sodium channels that eventually, if the stimuli is strong enough, lead to what we call an action potential. For a signal to become an action potential, the stimulus needs to bring the membrane potential around -50mV (which is called the threshold potential) by opening some sodium channels that let positively charged sodium ions enter the cell: it leads to the depolarization.
Another well known amino acid neurotransmitter is the gamma-aminobutyric (also called GABA) which have an inhibitory effect on brain synapses by increasing the membrane permeability to Cl- (you will see ions effect on the action potential in part 3).
When enough sodium channels are open, there is a retroaction cycle and even more sodium channels open (because these channels only open if the potential is positive enough and the more channels are open, the more positive the membrane gets). You can think of it like dominos.
Once the threshold potential is reached, the signal is an action potential (otherwise it just becomes a signal unable to create depolarization strong enough in other cells, decreases with the distance and ceases) and the retroactive loop (domino effect) quickly brings the potential near ENa=+62mV (which is the equilibrium potential of Na+ for which there is no flux of this ion).
3) Repolarization:
However the membrane potential never reaches this equilibrium potential of Na+ because of two points:
– Na+ voltage-gated channels enter an “inactivated” state: the gate m remains opened but gates h are now closed. Then there is no more Na+ flow through the cell.
– K+ voltage-gated channels are now open as the potential is now really positive.
The consequence of that will be the massive flow of K+ from the cell to the extracellular space.
These events quickly bring the membrane potential close to the potential of EK+=-90mV (that is why this phase is called repolarization).
4) Hyperpolarization and refractory period:
This phase is only the consequence of the repolarization and occurs at the end of it when the membrane potential becomes more negative than its resting value.
That can be explained by the fact that there is no more Na+ flow, so K+ becomes the only type of ions that can influence the membrane potential. Then the membrane potential tends to -90mV but never reaches it as when the membrane potential becomes negative, K+ voltage gated channels are back to their closed state.
A really important point is the fact that because Na+ voltage channels remain closed during the repolarization/hyperpolarization phase, if a second stimulus arrives, it is not able to create an action potential unless its intensity is as much high than the previous one. This period is called the refractory period (it lasts for 1 to 2 ms for most of the neurons) and it is the main reason of why signals only go in one direction: if a signal arrives in the opposite direction of an action potential, it will not be able to go through the refractory period of the first one.
Here is a figure that sums up all the stages of the action potential creation:

We are now able to explain how the action potential is created but a question still remains: how is this action potential transferred from its initial neuron to other cells ?
b) The action potential propagation
Just as we said before, when the first action potential is created at the axon hillock, there is a flow of Na+ entering the cell. This depolarization creates an electric current near the region where it was created, a current which is strong enough to depolarize the area of the membrane that is next to it, and reaches the threshold (creating another action potential) and so on, the potential is spread across the axon.
Note that the newly created action potential has the same intensity and length than the previous one as the creation of a potential follows the all-or-none principle (meaning that if the starting intensity is not high enough, there will be no potential and if it is high enough, it will create a potential that will always go to a maximum of intensity around 30-50mV).

We will now speak about some mechanisms that optimize the potential propagation.
– The axon diameter has a great impact on an action potential. Indeed, here is the resistance’s formula for a wire (but it also works for a tube-like structure like axon):

We can clearly see with this formula that the resistance of an electrical current is proportional to 1/s, so the bigger the axon diameter is, the lower is the resistance and the better is the propagation of the potential!
– The electrical insulation is really important to improve the speed of the signal (as for vertebrate, axon diameter is not that great (mainly 20 um) and is provided by myelin shifts that we have seen in the first part of the chapter.
c) Synaptic potential and summation
Looking at the level of synapses, we can define two types of potential that are created as the result of the linkage between neurotransmitters (seen in the previous part) and membrane receptors:
– The excitatory postsynaptic potential (EPSP) is a depolarization created by specific neurotransmitters linkage to ionic channels that are only permeable to ions (Na+ for exemple). The resulting flows increase the membrane potential (more likely to create an action potential).
– The inhibitory postsynaptic potential (IPSP) is an hyperpolarization created by specific neurotransmitters linkage to ionic channels that are only permeable to ions (Cl– or K+ for exemple). The resulting flows decrease the membrane potential (less likely to create an action potential).
We will now deal with postsynaptic potential summation. Indeed, one single neuron can receive signals from multiple terminal branches of presynaptic neurons, thus these signals have to be summed in order to deliver a final response.
We distinguish two types of summation:
– Temporal summation is a phenomenon that occurs when two postsynaptic potentials from the same synapse arrive one after the other in a really small range of time (meaning that the first postsynaptic potential is not over when the second arrives).
– Spatial summation is a phenomenon that occurs when two or more postsynaptic potentials from two or more different synapses arrive at the same time.
Note that every type of postsynaptic potential (inhibitory or excitatory) can be summed following a certain logic that is:
– If two excitatory potentials are summed, the final one has a higher intensity (better chances to create an action potential).
– If two inhibitory potentials are summed, the final one has a lower intensity (no chances to create an action potential.
– If one excitatory and one inhibitory potentials are summed, the final one intensity will be the difference between the intensity of the excitatory one and the intensity of the inhibitory one (no chances to create an action potential).


Une réflexion sur “II. Communication between neurons”